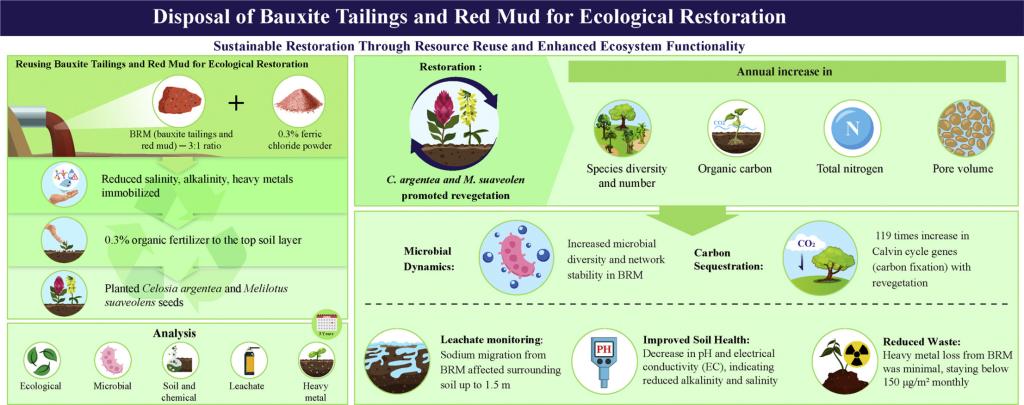
导读
铝土矿尾矿泥和赤泥通常作为铝土矿工业固体废物堆放,再利用潜力有限。本研究探究了如何将这两种固废进行资源再利用:作为类土壤基质(BRM)应用于铝土矿采空区的植被恢复。在为期三年的田间试验过程中,两种先锋植物——青葙(Celosia argentea)和草木樨(Melilotus suaveolens)成功地在场地建立了初始植被并促进了植被恢复,而植被恢复显著提高了BRM微生物多样性和网络稳定性(p< 0.05)。此外,微生物碳、氮固定基因丰度在BRM中也有所增加。其中,卡尔文循环相关基因丰度增加了119倍。随着生态处置,BRM土壤有机碳、全氮和孔隙体积呈逐年增加趋势,盐碱状况向自然土壤转变。BRM的三维孔隙结构显示其上层出现直径为400 ~ 800 μm的大孔隙。这些结果反映了BRM可持续性的生态能力。此外,场地淋溶液监测结果表明,BRM中有毒元素和盐分的环境迁移风险是可控的。研究结果展示了将工业固废转化为类土壤资源的潜力,提出了铝土矿固体废物可持续管理的策略。
论文ID
原名:Ecological disposal of bauxite tailings and red mud: A sustainable strategy for bauxite industrial waste reuse
译名:铝土矿尾矿泥和赤泥的协同生态处置:铝土矿工业固体废物再利用的可持续策略
期刊:Resources, Conservation and Recycling
IF:11.2
发表时间:2025 .3
通讯作者:张学洪刘杰
通讯作者单位:桂林理工大学
DOI号:10.1016/j.resconrec.2025.108259
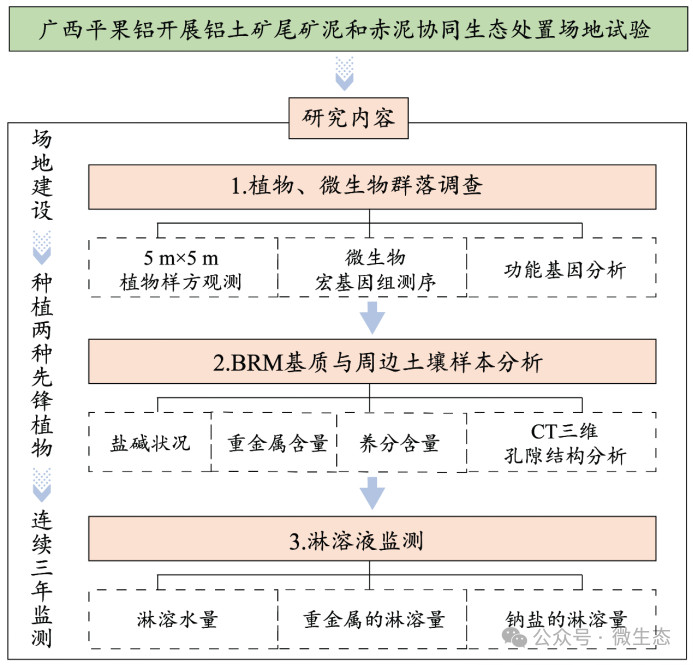
实验设计
结果
1.植物群落
经过3年的生态处置,BRM处置区内形成了初步的植物群落(图1)。BRM处置区内植被覆盖度从2021年的26.0%逐年增加到2023年的92.1%(p< 0.05)。与2021年相比,2023年地上生物量显著增加了168%。此外,Margalef和Pielou指数也显著增加,表明BRM处置区物种丰富度显著增加。
植物调查显示,从2021年到2023年,BRM处置区内的物种数量显著增加。在2021年,除人工种植的两种植物(青葙和草木樨)外,只有6种植物入侵并在处置区内存活。而2023年处置区内共发现13科26种,其中禾科10种。人工种植的种属中青葙和草木樨的价值最高,表明其在植物群落重建中发挥了重要作用。2023年,三叶鬼针草(Bidens pilosa)、白背黄花稔(Sida rhombifolia)和狗牙根(Cynodon dactylon)的重要值分别为7.36、5.40和5.40,表明这3种植物具有成为优势物种的潜力。2023年1棵桉树(Eucalyptus robusta)和5棵灌木成功地在BRM处置区定居,表明植物群落结构的复杂性可能会增加。

图1. 2021年至2023年,对铝土矿尾矿泥和赤泥类土基质(BRM)处置区进行植被重建。(a)处置区植被图片;(b)植被覆盖率;(c)物种数量;(d)Shannon多样性指数;(e)地上生物量;(f)Margalef丰富度指数;(g)Pielou指数。星号表示根据 t检验,p< 0.05;p< 0.01;p< 0.001时存在显著差异; ns表示无显著性。
2. 微生物群落
生态处理后,BRM中微生物Shannon多样性指数呈波动上升趋势(图2)。2023年10月BRM微生物多样性较2021年4月显著增加14.9% (p< 0.05)。主坐标分析(PCoA)表明,2021年和2023年BRM微生物群落的聚集性较高,群内变异较小。相比之下,微生物群落的聚集性较低2022年群落聚集度较低,表明群落变异性相对较高。2021年后,BRM微生物群落结构发生显著变化(p< 0.05)。在门水平上,BRM中的优势微生物包括15种细菌,4种真菌和3种古细菌。2021年4月以后,优势真菌和古菌的丰度分别由0.07%和0.10%增加到0.11% ~ 0.62%和0.43 ~ 2.05%。此外,BRM中优势门假单胞菌门(细菌)丰度从81.4%下降到38.2% ~ 62.6%,放线菌门丰度从10.8%上升到12.8%和25.0%。到2023年10月,BRM中的优势属假单胞菌的丰度从23.3%下降到1.86%,而慢生根瘤菌和硝化螺旋菌的丰度分别增加了2.53倍和104倍(图S4)。参与碳和氮循环的微生物丰度的增加也导致了BRM中功能基因丰度的变化。从2021年4月到2023年10月,碳和氮固定基因的丰度分别从5.98和0.00增加到37.5和15.9(图S5)。为了评估生态处置对BRM中微生物相互作用的影响,我们构建了微生物生态网络(图S6)。随着生态处理时间的增加,BRM中微生物网络的规模(总节点数)逐渐扩大。值得注意的是,到2023年,BRM网络的复杂性将显著提高增加,节点、链路和平均程度的数量显著增加。鉴定出BRM微生物网络中具有重要作用的15个关键物种(6个模块枢纽和9个连接器)。有趣的是,随着处理时间的延长,这些关键物种的丰度发生了变化。2021年,BRM处置区内所有关键物种的丰度均较低。但从2022年到2023年,不同关键物种的丰度显著增加。
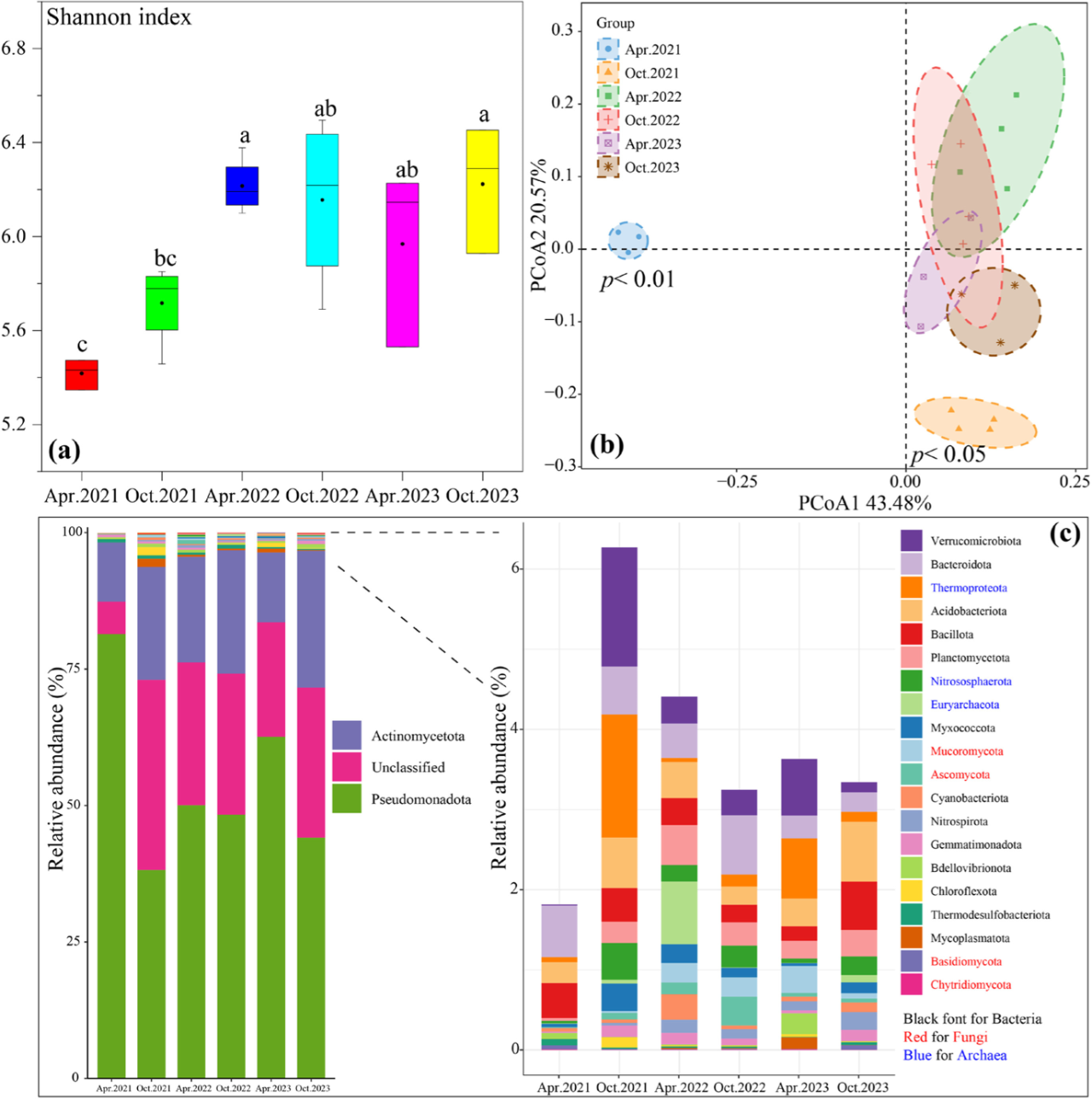
图2. 2021年至2023年BRM中微生物群落的演替。(a)Shannon多样性指数;(b)主坐标分析法(PCoA)分析了微生物群落结构;(c)优势微生物的相对丰度(门水平,相对丰度> 0. 05%)。
3. BRM处置对周边环境的影响
2021年5月至2022年4月处置区每月Na和重金属浸出量如图5所示。处置区Na浸出主要发生在5 ~ 10月,月平均浸出量为51.2 mg/m²,8月达到108 mg/m²的峰值。这一趋势总体上与渗滤液体积的变化一致。2022年2 - 4月,虽然处置区内的渗滤液量达到606 ~ 841 mL/m²,但Na浸出量仅为3.44 ~ 12.9 mg/m²。此外,周边土地的Na淋滤量相对较低,在2021年5月达到18.2 mg/m²的峰值后,每月均保持在5 mg/m²以下。
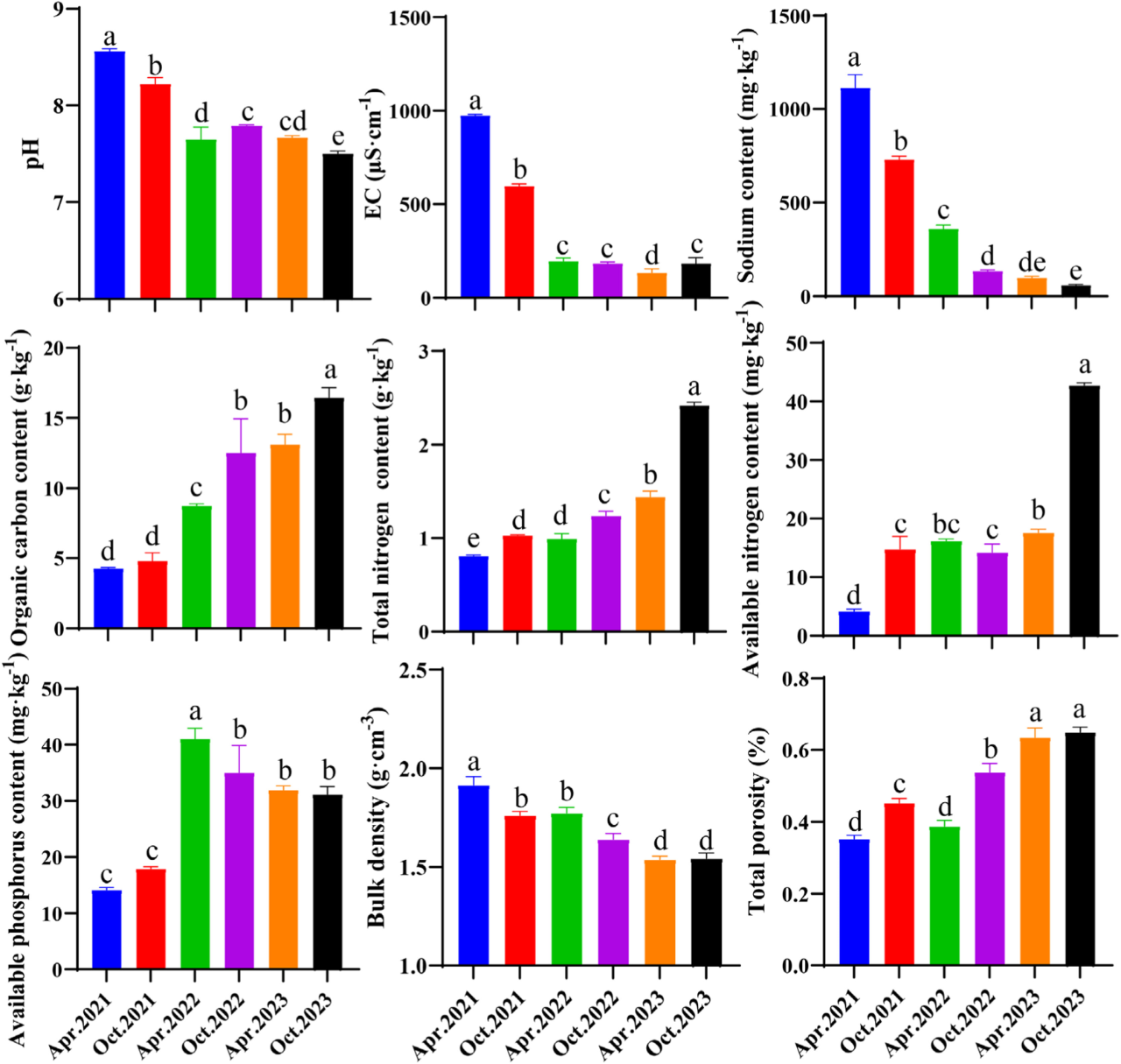
图3. 采用生态处理开发的BRM基质的理化性质。不同小写字母代表不同处理时间均值之间的显著差异(最小显著差异[LSD],p< 0.05)。

图4. BRM基质中孔结构的变化。(a)不同处理时间下BRM中孔的三维分布。(B)BRM中不同直径孔的体积。

图5. BRM 处置区和周围 3 m 以内的土地每月浸出的 Na 和重金属量。
随着BRM生态处置时间的延长,周边1 m范围内土壤盐度和碱度呈现先升高后降低的趋势(图6),到2022年4月,pH达到峰值EC值在2022年10月达到峰值122 μS/cm。但到2023年10月,pH和EC值分别降至7.31和83.7 μS/cm。在BRM处置的3年期间,周边3m范围内的土壤pH和EC值没有显著变化。另外,与2021年4月相比,到2022年10月,周围土壤Na含量下降了44.1-52.1%。
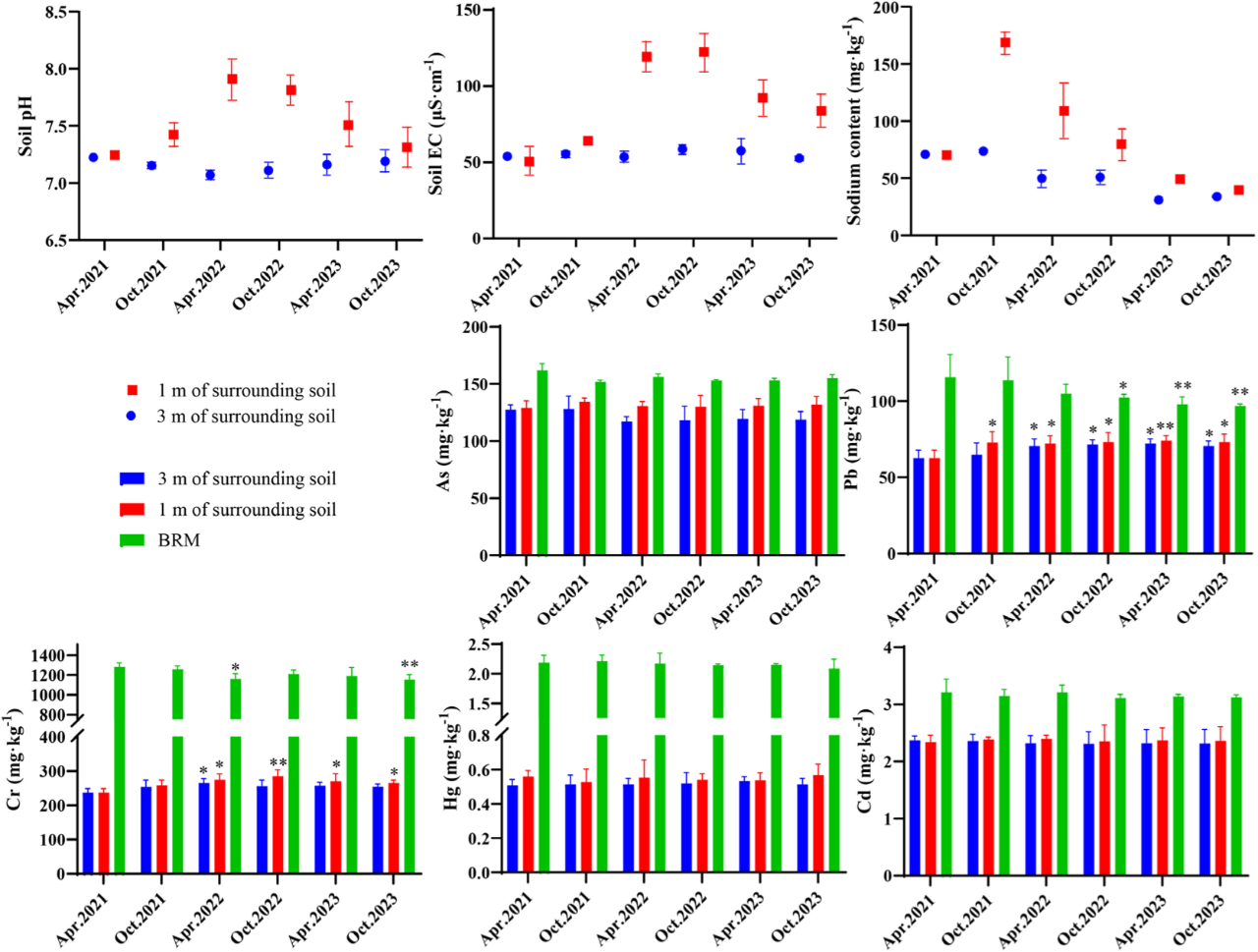
图6. BRM 生态处置对周围土壤盐度和重金属含量的影响。采用独立样本 t 检验比较 BRM 或土壤样本2021年4月与其他处置时间之间的显著差异,p< 0.05;p< 0.01;p< 0.001。
作为两种铝土矿固体废物的混合物,BRM中这五种重金属的含量高于周围土壤。BRM中Cr、As和Pb含量分别为1280、162和116 mg/kg(图6)。BRM生态处理区的每月重金属浸出量保持在150 μg/m²以下。而周边土地5月份的As浸出量为163 μg/m²(图5)。处置区内重金属的浸出主要发生在8 - 10月。处置区Pb和Cr的淋溶水平高于周边地区。10月,处置区内Pb浸出量为121 μg/m²,周边地区为49.8 μg/m²。8月处置区Cr浸出量为32.4 μg/m²,周边土地Cr浸出量为2.71 μg/m²。在生态处置过程中,BRM及周边土地As、Hg、Cd含量均无显著变化(p> 0.05)。而到2023年10月,BRM中Pb和Cr浓度分别显著下降了16.2%和9.96% (p< 0.05)。周边1 m和3 m范围内的土壤Pb浓度显著增加,分别从2021年4月的62.7 mg/kg和62.3 mg/kg增加到2023年10月的73.2 mg/kg和70.5 mg/kg。与此同时,土壤中Cr浓度呈先上升后下降的趋势,到2023年10月与2021年4月相比,距周围土壤1 m和3 m范围内的Cr浓度分别上升了11.9%和6.98%。
讨论
1 生态处置区的生态修复
本研究在铝土矿尾矿泥和赤泥协同生态处置区内成功种植了两种先锋植物——青葙和草木樨。这有助于BRM处置区内植物群落的建立,植被覆盖度、物种数量和多样性指数每年都在增加(图1)。在2021年10月的植被调查中,成功建立了6个新物种(表S1)。这说明处置区具有一定的自主恢复能力,植被结构更加多样化。值得注意的是,青葙和草木樨的花期在5月至9月,吸引了大量的蜜蜂,从而促进了种子库的引入。在尾矿库和赤泥场建立稳定的植物群落需要很长时间。本研究经过3年的生态BRM处理,在处置区内共发现13科26种植物。虽然植物群落以草本植物为主,但也有1种乔木(桉树)和5种灌木。这表明BRM处置区生态演替向更稳定的阶段发展。这可以部分归因于两种先锋植物为后期演替物种创造了生态位机会。例如,先锋植物为种子落地生根提供了有利条件,如凋落物分解的养分输入(图3)和根生长改善的孔隙结构(图4)。此外,有利的当地气候,包括充足的降雨和温度(图S1),进一步支持了植物在实验场地的建立和生长。
微生物是生态系统不可缺少的组成部分,在生物地球化学过程中发挥关键作用。然而,采矿活动严重破坏了固体废物中的微生物群落。在2021 - 2022年间,BRM微生物群落的Shannon多样性指数增加,群落组成发生了实质性变化。这可能是由于植物根系形成了大孔隙,为微生物的快速增殖提供了栖息地(图4)。BRM微生物群落多样性指数呈波动变化,反映了微生物演替的“螺旋式上升”模式。尽管微生物多样性没有显著增加,但微生物网络的复杂性显著增加(图S6),表明BRM中微生物之间的相互作用增强。此外,微生物网络中关键物种的丰度随着处理时间的增加而发生变化。例如,连接器链霉菌(Streptomycescavourensis)和模块枢纽分枝杆菌(Mycolicibacterium neoaurum)在2022年4月的丰度较高,它们以对盐度和碱度的耐受性以及有机酸的分泌而闻名。此外与蚂蚁共生定殖的Pseudonocardia broussonetiae作为模块化枢纽,在2022年丰度显著增加,这一年我们在研究地点发现了许多蚂蚁巢穴(图S2)。到2023年10月,模块枢纽Rhodo -细菌科细菌QY30,一种以其在碳水化合物降解和有氧反硝化中的作用而闻名的细菌,丰度显著增加。这表明,不同演替阶段需要不同的关键微生物参与者来实现不同的生态功能。这也反映了BRM地下生态需求的转变,从盐度和碱度调节到养分积累。重要的是植物和微生物群落的恢复也证明了处置区生态恢复的成功,并进一步减少了铝土矿开采对环境的影响。
2.从BRM到类土基质的演变
BRM的土壤形成对处置区的生态可持续性至关重要。BRM与天然土壤的主要区别在于盐度、碱度、孔隙度和营养成分,这些土壤是由铝生产过程(如湿磨和氢氧化钠消化)产生的。
盐度和碱度:铝土矿尾矿泥的加入稀释了赤泥的高盐度和碱度。此外,添加氯化铁会产生H+,其中和碱度并进一步稳定BRM中的碱性矿物。BRM的初始pH值为8.56,EC值为974 μS/cm(图3),均符合标准《绿化种植土壤》(CJ/T 340–2016)。生态处置一年后,BRM的pH和EC 显著降低,随后趋于稳定。这归因于先锋植物草木樨对Na的强大积累能力和降雨的浸出效应,这阻止了BRM处理区的表面盐碱化。XRD图谱表明碱性矿物Cancrite的衍射峰强度略有降低,对应于BRM中碱度的降低(图 S7)。此外,水化石榴子石(Hydrogarnet)和钙锆榴石(Kimzeyite)次生矿物的出现表明BRM向土壤的转变。
孔隙度:在粘土中形成稳定的大孔隙是具有挑战性的。Yan等(2024)报道了盐碱土壤中钠盐的浸出会破坏孔隙结构。然而经过3年的生态处理,BRM的孔隙体积增加了196%,这是一个显著的变化(图4)。这可能是由于植物的根际活动(图S2)。而BRM颗粒在根通道周围的排列形成孔壁,在根腐烂后保持稳定,形成致密的孔。CT扫描显示,到2023年10月,BRM上层出现了直径在400 ~ 800 μm之间的孔隙。这可能是由于地表凋落物和根状沉积物的分解,导致有机聚合物的产生,这些有机聚合物结合粘土颗粒形成稳定的孔隙。
营养:植物和微生物群落的恢复促进了基质中营养物质的积累。在三年的处理过程中,由于成功的植被恢复策略,BRM中的有机碳、总氮和有效磷含量稳步增加(图3)。首先,这两种先锋植物是一年生植物,在冬季死亡和枯萎,有助于有机碳的输入。豆科植物如M. suaveolens和Sesbania canna(表S1),被慢生根瘤菌等微生物寄生,形成根瘤固定氮(图S2)。根瘤菌丰度的增加和固氮基因(nifKDH)的表达进一步促进了BRM中氮的积累(图5)。此外,与2021年4月相比,BRM中碳固定相关基因的丰度显著增加,2023年10月增加了5.27倍(p< 0.05)。三种碳固定途径分别为在BRM中检测到:Calvin-Benson-Bassham (CBB)循环、rTCA循环和3-羟基丙酸双循环(3-HB)。2021-2022年期间,碳固定途径主要为rTCA循环和3-HB途径。到2023年,CBB循环是主要的碳固定途径。不同的代谢途径在碳固定过程中表现出不同的能量需求。在缺乏能量的栖息地,参与碳固定的微生物更喜欢低能量需求的途径,如rTCA循环(需要两个ATP分子)和3-HB途径(需要五个ATP分子)。然而,CBB循环需要9个ATP分子。值得注意的是从2021年4月到2023年10月,BRM中CBB循环(prkB)的丰度显著增加,从0.25增加到30.1(图S5)。这表明在BRM中有更多的能量可用。综上所述,经过三年多的生态修复,BRM 稳步向土壤转化,显著提高了处置区域的生态可持续性。盐度、孔隙度和养分积累的改善表明了生态处置的成功,以及BRM 在铝土矿场大规模就地处置和植被恢复方面的潜力。这些结果表明,BRM能够支持可持续的类土壤生态系统,其中植物和微生物群落都能循环利用关键元素,并随着时间的推移增强生态可持续性。
3.铝土矿固废协同生态处置技术的安全性
工业固体废物资源化利用对保护环境、节约资源、促进可持续发展具有重要意义。工业固体废物的安全处置是实现资源化利用的重要前提。BRM受到采矿活动和该地区地质背景的严重影响,导致Na和重金属含量高。BRM中的Na含量明显高于周围土壤(图4和图7),Na可能由于雨水淋滤而迁移到邻近土壤。60 ~ 100 cm深度的土壤监测表明,随着时间的推移,离处置场地1 m的土壤中Na含量先升高后降低,pH值和EC值也呈现出类似的变化趋势。然而,在距离处置区3 m的土壤中没有观察到明显的变化。这表明Na从BRM的横向迁移被限制在3米半径内。Pietrzyk-Sokulska和Kulczycka(2015)调查了波兰南部的戈尔卡赤泥堆,发现即使在30年后,残留的赤泥仍在继续增加半径20米范围内附近土壤的盐度。说明赤泥与铝土矿尾矿泥协同生态处置后,BRM的环境影响半径显著减小,潜在的环境风险得到缓解。在雨季,每月从BRM处理区域的浸出量为28.1至108 mg/m²(图5)。估计表明,每月约有0.1-2%的Na从BRM中浸出。从2023年4月到10月,BRM表层Na含量显著下降34.6% (p< 0.05)。这种减少归因于植被对Na的吸收和表面BRM孔隙结构的改善,从而促进了淋滤。此外,由于处置区的压实作用,BRM致密的中间层限制了Na的进一步向下迁移。此外,BRM处置区内的Na浸出浓度低于《中国地下水水质标准》(GBT14848-2017)规定的限值(表S2)。
具有高比表面积的BRM颗粒和氯化铁水解形成的Fe (OH)₃絮凝体对重金属离子具有很强的吸附能力。在2021 - 2022年期间,BRM生态处置区内重金属的月淋溶量保持在150 μg/m²以下(图5)。然而,随着BRM pH值的降低(图3),某些重金属(如Pb、Cd和Cr)的迁移率在5月9日下增加。值得注意的是,BRM中Cr和Pb的浓度分别下降了16.2%和9.96%(图6,p< 0.05)。周边土壤Cr、Pb浓度显著升高,但未超过《中国农用地土壤污染风险控制标准》(GB15618-2018)。研究结果表明,在生态处置区内,BRM中Na和重金属的环境风险是可控的。然而,Pb和Cr在周围土壤中的长期迁移需要持续监测。
4.铝土矿固废协同生态处置技术的可行性
矿泥库和赤泥堆场的建设、管理和维护费用昂贵。以本研究的广西平果铝土矿为例,投资2亿多建设的尾矿库只能使用5年,每年的处置费用超过5000万。工业废弃物的堆积性严重限制了铝土矿工业的可持续发展潜力。此外,世界范围内大面积矿山景观的植被恢复需要通过挖掘自然景观来获取大量的自然土壤,这不是一种可持续的资源管理模式。我们的研究结果展示了一种范式转换的解决方案。通过生态工程将铝土矿尾矿泥和赤泥(BRM)转化为替代土壤,我们消除了对天然表土的需求——考虑到78%的矿区缺乏可获取的表土储量,这是一个至关重要的优势。与传统的土壤进口方法相比,这种方法降低至大约22元/吨BRM的植被恢复成本(表S3),实现了成本节约。至关重要的是,相邻铝土矿废物(铝土矿尾矿泥和赤泥)的协同处置最大限度地减少了长途运输。通过将废物转化为替代土壤,这一过程代表了回收工业废物和减少铝土矿工业环境足迹的实用方法。此外,这种生态处理策略虽然没有充分利用工业固体废物中的剩余资源价值,但可以迅速缓解环境压力,降低管理成本。本研究的结果将有助于我们更好地了解工业固体废物的生态处理,这对铝土矿开采的可持续发展至关重要。
结论
在这项研究中,我们展示了两种铝土矿工业固体废物如何作为类土壤基质进行生态处置以实现资源再利用。通过将废物转化为有价值的生态资产来减少铝土矿开采的环境足迹。在BRM上成功生长的青葙和草木樨不仅促进了植被恢复,而且有助于基质中养分积累。此外,微生物多样性和稳定性的增加进一步反映了BRM支持可持续类土生态系统的能力。有毒元素(Cd、As、Pb、Hg、Cr)的损失量较低(每月< 150 μg/m²),表明了铝土矿协同生态处置的安全性。将BRM作为类土壤基质应用于铝土矿采空区生态修复,我们减少了对客土的需求,限制了土地资源的开采,促进了工业废物的可持续利用,有助于实现环境保护和固废缩量化的目标。
原文链接:https://doi.org/10.1016/j.resconrec.2025.108259
